Da bi lahko celovito govorili o hemoglobinu (Hb), je koristno najprej obravnavati mioglobin (Mb), ki je zelo podoben hemoglobinu, vendar je veliko enostavnejši. Med hemoglobinom in mioglobinom obstajajo strogi sorodstveni odnosi: oba sta konjugirana proteina in njihova protetična skupina (neproteinski del) je heme skupina.
Mioglobin je globularni protein, sestavljen iz ene verige s približno sto petdesetimi aminokislinami (odvisno od organizma) in njegova molekulska masa je približno 18 Kd.
Kot je omenjeno, je opremljen s heme skupino, ki je vstavljena v hidrofobni (ali lipofilni) del proteina, sestavljen iz zloženk, ki jih lahko pripišemo α-helix strukturam vlaknastih proteinov.
Mioglobin je sestavljen predvsem iz segmentov α-spiral, prisotnih v številu osem in sestoji, skoraj izključno, iz nepolarnih ostankov (levcin, valin, metionin in fenilalanin), medtem ko so polarni ostanki praktično odsotni (asparaginska kislina, glutaminska kislina, lizin in arginin); edini polarni ostanki sta dva histidina, ki imata ključno vlogo pri napadu kisika na heme skupino.
Heme skupina je kromoforska skupina (absorbira se v vidnem) in je funkcionalna skupina mioglobina.
Glejte tudi: glikirani hemoglobin - hemoglobin v urinu
Malo kemije
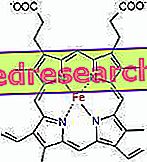
Povezava med protoporfirinom in železom je tipična vez tako imenovanih koordinacijskih spojin, ki so kemijske spojine, v katerih osrednji atom (ali ion) tvori vezi z drugimi kemičnimi vrstami v številu, ki je višja od njegove oksidacijske številke (električni naboj). V primeru hema so te vezi reverzibilne in šibke.
Koordinacijska številka (število koordinacijskih vezi) železa je šest: okoli železa je lahko šest molekul, ki si delijo vezne elektrone.
Za tvorbo koordinacijske spojine potrebujemo dve orbitali s pravilno orientacijo: osebo, ki lahko "kupi" elektrone, druga pa jih lahko podari.
V hemu železo tvori štiri ravninske vezi s štirimi atomi dušika, ki so v središču proto-porfirinskega obroča in peto vez z proksimalnim histidinskim dušikom; železo ima šesto povezavo brez koordinacije in se lahko veže na kisik.
Ko je železo v obliki prostega iona, imajo vse njegove orbite tipa d enako energijo; v mioglobinu je železov ion vezan na protoporfirin in histidin: te vrste magnetno motijo orbitale železa; obseg motenj bo različen za različne orbitale glede na njihovo prostorsko usmerjenost in vznemirljive vrste. Ker se mora celotna energija orbitalov vrniti v konstanto, motnje povzročajo energetsko ločitev med različnimi orbitali: energija, ki jo pridobijo z nekaterimi orbitali, je enaka energiji, ki jo izgubijo druge.
Če ločitev, ki se pojavi med orbitali, ni zelo velika, je zaželena elektronska ureditev z visokim vrtenjem: elektroni za vezanje se poskušajo vzporedno vrteti v več možnih pod nivojih (največja množina); če pa je motnja zelo močna in je velika razdalja med orbitali, je lahko bolj primerno, da se povežejo vezni elektroni v nižjih energijskih orbitalah (nizek spin).
Ko se železo veže na kisik, molekula prevzame nizek položaj spina, medtem ko ima železo šesto povezavo brez koordinacije, molekula ima visoko vrtenje.
S to razliko v spinu, s spektralno analizo mioglobina, lahko razumemo, ali je vezan kisik (MbO2) ali ne (Mb).
Mioglobin je beljakovina, značilna za mišice (vendar ne le v mišicah).
Mioglobin se ekstrahira iz kita, v katerem je prisoten v velikih količinah in nato prečiščen.
Kiti pri dihanju kot človeška bitja: pljuča morajo absorbirati zrak skozi dihalni proces; kita za spermo mora nositi čim več kisika v mišicah, ki so sposobne kopičiti kisik, tako da se vežejo na prisotni mioglobin; kisik se nato sprosti počasi, ko se potopi kitov, ker je za presnovo potreben kisik: večja je količina kisika, ki jo lahko kobila absorbirajo in več kisika je na voljo med potopom.

Da bi bilo meso bolj ali manj rdeče, je vsebnost hemoproteinov (to je heme, ki naredi meso rdeče).
Hemoglobin ima številne strukturne analogije z mioglobinom in lahko reverzibilno veže molekularni kisik; medtem ko je mioglobin omejen na mišice in periferna tkiva na splošno, se hemoglobin nahaja v eritrocitih ali rdečih krvnih celicah (to so psevdo celice, tj. niso prave celice), ki predstavljajo 40% krvi.
Hemoglobin je tetrameter, kar pomeni, da ga sestavljajo štiri polipeptidne verige, vsaka s heme skupino in dve po dve (pri človeku sta dve alfa verigi in dve beta verigi).
Glavna funkcija hemoglobina je transport kisika ; Druga funkcija krvi, pri kateri sodeluje hemoglobin, je transport snovi v tkiva.
Na poti od pljuč (bogatih s kisikom) do tkiv, hemoglobin prenaša kisik (hkrati pa tudi druge snovi dosežejo tkiva), medtem ko na povratni poti nosi s seboj zbrane odpadke iz tkiv, zlasti ogljikov dioksid, ki nastane v presnovi.
V razvoju človeka obstajajo geni, ki se izražajo le za določeno časovno obdobje; zato imamo različne hemoglobine: fetalni, zarodni, odraslega človeka.
Verige, ki sestavljajo te različne hemoglobine, imajo različne strukture, vendar so podobne funkcije, ki jih opravljajo, bolj ali manj enake.
Razlaga prisotnosti več različnih verig je naslednja: med evolucijskim procesom organizmov se je razvil tudi hemoglobin, specializiran za transport kisika iz območij, ki so bogata na pomanjkljivih področjih. Na začetku evolucijske verige je hemoglobin prenašal kisik v majhne organizme; Med evolucijo so organizmi dosegli večje dimenzije, zato se je hemoglobin spremenil, da bi lahko prenašal kisik na območja, ki so bolj oddaljena od točke, ko je bil bogat; V ta namen so bile med evolucijskim procesom kodificirane nove strukture verig, ki tvorijo hemoglobin.
Mioglobin veže kisik tudi pri skromnih obremenitvah; v perifernih tkivih je pritisk (PO2) okoli 30 mmHg: pri tem tlaku mioglobin ne sprosti kisika, zato bi bil neučinkovit kot kisikov nosilec. Po drugi strani pa ima hemoglobin bolj elastično vedenje: veže kisik pri visokih tlakih in ga sprosti, ko se tlak zmanjša.
Ko je beljakovina funkcionalno aktivna, lahko nekoliko spremeni svojo obliko; na primer, kisikova mioglobin ima drugačno obliko od neogljikovega mioglobina in ta mutacija ne vpliva na sosednje.
Diskurz je drugačen v primeru povezanih beljakovin, kot je hemoglobin: ko je veriga oksidirana, se inducira, da spremeni svojo obliko, vendar je ta modifikacija tridimenzionalna, zato so prizadete tudi druge verige tetrametra. Dejstvo, da so verige med seboj povezane, vodi k razmišljanju, da sprememba enega vpliva na druge sosednje, čeprav v različnem obsegu; ko je veriga oksidirana, druge verige tetrametra prevzamejo "manj sovražni odnos" do kisika: težava, s katero se verige oksigenati zmanjšujejo, ko se verige zraven, oksidirajo same. Isti argument velja za deoksigenacijo.
Kvartarna struktura deoksihemoglobina se imenuje T (napeta) oblika, medtem ko se oksihemoglobin imenuje oblika R (sproščena); v raztegnjenem stanju obstaja vrsta precej močnih elektrostatičnih interakcij med kislinskimi aminokislinami in osnovnimi aminokislinami, ki vodijo v togo strukturo deoksihemoglobina (zato je "napeta oblika"), medtem ko je, ko je vezan kisik, entiteta teh interakcije se zmanjšajo (zato "sproščena oblika"). Nadalje, v odsotnosti kisika, je histidinski naboj (glej strukturo) stabiliziran z nasprotnim nabojem asparaginske kisline, medtem ko je v prisotnosti kisika težnja, da protein izgubi proton; vse to pomeni, da je kisikov hemoglobin močnejša kislina kot deoksigenirana hemoglobija: bohrov učinek .
Glede na pH se skupina heme bolj ali manj enostavno veže na kisik: v kislem okolju hemoglobin lažje sprošča kisik (napeta oblika je stabilna), v osnovnem okolju pa je vez s kisikom močnejši.

Vsak hemoglobin sprosti 0, 7 protonov na vhodni mol kisika (O2).
Bohrov učinek omogoča hemoglobinu, da izboljša svojo sposobnost prenašanja kisika.
Hemoglobin, ki potuje od pljuč do tkiv, se mora uravnotežiti glede na tlak, pH in temperaturo.
Vidimo učinek temperature .
Temperatura v pljučnih alveolah je za 1-1, 5 ° C nižja od zunanje temperature, v mišicah pa je temperatura okoli 36, 5-37 ° C; ko se temperatura zviša, se faktor nasičenosti zmanjša (z istim tlakom): to se zgodi zato, ker se kinetična energija povečuje in disociacija je prednost.

Obstajajo tudi drugi dejavniki, ki lahko vplivajo na sposobnost hemoglobina, da se veže na kisik, med katerimi je koncentracija 2, 3-bisfosfoglicerata .
2, 3-bisfosfoglicerat je presnovno sredstvo, prisotno v eritrocitih v koncentraciji 4-5 mM (v nobenem drugem delu telesa ni prisotno v tako visoki koncentraciji).
Pri fiziološkem pH se 2, 3-bisfosfoglicerat deprotonira in ima na njem pet negativnih nabojev; gre za zagozditev med dvema beta verigama hemoglobina, ker imajo te verige visoko koncentracijo pozitivnih nabojev. Elektrostatične interakcije med beta verigami in bisfosfogliceratnimi verigami dajejo sistemu določeno togost: dobljena je napeta struktura, ki ima majhno afiniteto za kisik; Med oksigenacijo se nato izloči 2, 3-bisfosfoglicerat.
V eritrocitih obstaja posebna naprava, ki pretvori 1, 3-bisfosfoglicerat (ki ga proizvaja presnova) v 2, 3-bisfosfoglicerat, tako da doseže koncentracijo 4-5 mM in je zato hemoglobin sposoben izmenjevati kisika v tkivih.
Hemoglobin, ki prispe v tkivo, je v sproščenem stanju (vezan na kisik), vendar je v bližini tkiva karboksiliran in prehaja v raztegnjeno stanje: beljakovina v tem stanju ima nižjo tendenco, da se veže s kisikom, v primerjavi s sproščenim stanjem, zato hemoglobin sprosti kisik v tkivo; poleg tega zaradi reakcije med vodo in ogljikovim dioksidom nastajajo ioni H +, zato je zaradi bohrjevega učinka še več kisika.
Ogljikov dioksid se razprši v eritrocit, ki prehaja skozi plazemsko membrano; glede na to, da eritrociti tvorijo približno 40% krvi, lahko pričakujemo, da le 40% ogljikovega dioksida, ki difundira iz tkiv, vstopi vanje, v resnici 90% ogljikovega dioksida vstopi v rdeče celice, ker vsebujejo encim, ki pretvarja. ogljikov dioksid v ogljikovi kislini sledi, da je stacionarna koncentracija ogljikovega dioksida v eritrocitih nizka, zato je hitrost vstopa visoka.

Naslednji pojav, ki se pojavi, ko eritrocit doseže tkivo, je naslednji: z gradientom HCO3- (derivat ogljikovega dioksida) izhaja iz eritrocita in, da bi izravnali izhod negativnega naboja, imamo vstop kloridov, ki določajo povečanje osmotskega tlaka: za uravnoteženje te variacije je tudi vstop vode, ki povzroča otekanje eritrocitov (učinek HAMBURGER). Nasprotni pojav se pojavi, ko eritrocit doseže pljučne alveole: pride do deflacije eritrocitov (učinek HALDANE). Tako so venski eritrociti (neposredno v pljuča) okrogli kot arterijski.


